
Содержание
Группы сцепления
В начале двадцатого века Томас Хант Морган (1866–1945 гг.) пытался проверить, как действует закон Менделя независимого наследования признаков у животных. Т. X. Морган – американский зоолог. Работал в Колумбийском университете и Калифорнийском технологическом институте. На основе исследований мутаций у дрозофилы, проведенных вместе с Г. Дж. Меллером, А. Стертевантом и К. Бриджесом, обосновал представление о материальных носителях наследственности. За сформулированную на основе этих исследований хромосомную теорию наследственности награжден Нобелевской премией (1933 г.).
Исследования проводились на плодовой мушке дрозофиле (Drosophila melanogaster). Преимущества избранного исследовательского материала:
– имеет небольшое количество хромосом – четыре пары;
– легко содержать;
– имеют значительную плодовитость;
– быстрая смена поколений – каждые полторы-две недели.
Количество генов больше количества хромосом. То есть в каждой хромосоме – большее количество генов. Гены, лежащие в одной хромосоме, называют сцепленными. Все гены, лежащие в одной хромосоме, образуют группу сцепления. Они попадают в одну гамету и наследуются вместе. Наличие групп сцепления установлено для всех генетически изученных организмов. Число групп сцепления у организмов равно числу пар хромосом и соответствует числу хромосом в гаплоидном наборе. Например, у кукурузы их 10, у дрозофилы – 4, у человека – 23.
Для генетически менее изученных видов пока что известны не все гены, поэтому число групп сцепления в них несколько меньше числа пар хромосом. Например, у кроликов 22 пары хромосом, а групп сцепления известно лишь 11. У вирусов и бактерий все гены относятся к одной группе сцепления.
Таким образом, из-за сцепления генов они в одной хромосоме наследуются все вместе, тогда как гены, расположенные в разных хромосомах, могут независимо комбинироваться в процессе мейоза по законам случайного распределения.
Кроссинговер
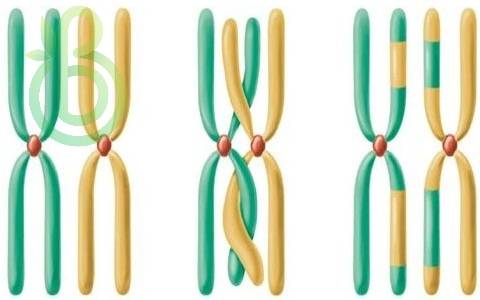
В результате исследований оказалось, что гены, которые находятся в одной хромосоме, сцеплены не абсолютно. Во время мейоза в профазе I при конъюгации гомологичные хромосомы обмениваются участками между хроматидами – происходит скрещивание или кроссинговер. Кроссинговер может происходить в любом участке хромосом, даже в нескольких участках. Чем дальше расположены участки, тем больше вероятность кроссинговера. Кроссинговер является обязательным процессом и происходит в каждой паре гомологичных хромосом. Во время такого обмена происходит рекомбинация генов, возрастает многообразие гамет с разным объединением генов.
Все гаметы, которые имеют хромосому, измененную благодаря кроссинговеру, называются кроссоверными. Потомство, в образовании которого принимали участие кроссоверные гаметы, – кроссоверы.
Гаметы, которые несут хромосомы, в которых не произошло изменений, называют некроссоверными. Потомство, образовавшееся из таких гамет, – некроссоверы.
Процент кроссоверов к общему количеству организмов в потомстве называется частотой кроссинговера. Частота кроссинговера зависит не только от расстояния между генами. Для ряда локусов выявлено, что частота кроссинговера в мейозе увеличивается с возрастом.
Кроссинговер характерен для большинства растений и животных за исключением самки тутового шелкопряда и самца мушки дрозофилы.
Частота кроссинговера и величина сцепления генов
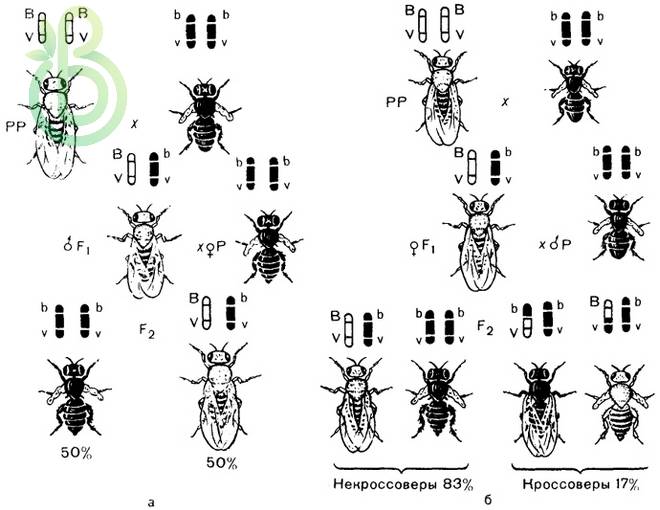
В случае независимого наследования генов гетерозиготные по двум признакам особи вследствие анализирующего скрещивания в потомстве проявляли расщепление в соотношении 1:1:1:1.
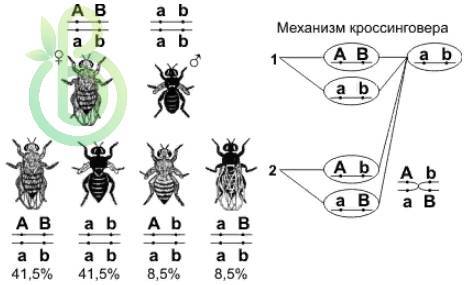
В опыте Моргана анализирующее скрещивание дало другие результаты. У дрозофилы гены, контролирующие окраску тела (серая – черная) и длину крыльев (длинные – короткие зачатки) локализованы в одной хромосоме. Серое тело и длинные крылья доминируют.
При скрещивании гомозиготной по рецессивным признакам самки с дигетерозиготным самцом происходит расщепление 1:1 по фенотипу.
В – серое тело, b – черное; V – длинные крылья, v – короткие:
Р: ♀ bv / / bv × ♂ BV / / bv
черное тело серое тело
короткие крылья длинные крылья
G: bv BV, bv
F: BV / / bv, bv / / bv
серое тело черное тело
длинные крылья короткие крылья
50 % 50 %
Гены, которые локализованы в одной хромосоме, передаются всегда вместе – такое сцепление называется полным. Дигетерозиготная особь в этом случае образует вместо четырех типов гамет лишь два.
У самца невозможен процесс конъюгации и кроссинговер, вследствие чего не образуются кроссоверные гаметы. Это можно объяснить отсутствием у самцов мушек дрозофил особого белкового комплекса, необходимого для обеспечения процесса конъюгации гомологичных хромосом.
При скрещивании дигетерозиготных самок дрозофил с гомозиготными по рецессивным генам самцами возникает расщепления в потомстве:
Р: ♀ BV / / bv × ♂ bv / / bv
серое тело черное тело
длинные крылья короткие крылья
G: BV, bv bv
Bv, bV
F: BV / / bv, bv / / bv Bv / / bv, bV / / bv
серое тело черное тело серое тело черное тело
длинные короткие короткие длинные
крылья крылья крылья крылья
41,5 % 41,5 % 8,5 % 8,5 %
У дигетерозиготной самки дрозофилы образуются четыре типа гамет, два из которых (BV, bv) – некроссоверные, а два других (Bv, bV) – кроссоверные, которые возникли вследствие кроссинговера.
В потомстве появились особи, которые отличаются по своим признакам от родительских форм (с серым телом, короткими крыльями и черным телом, длинными крыльями) – имеют рекомбинантные фенотипы.
Два или больше генов являются сцепленными неполностью, если потомки с новыми генными комбинациями (рекомбинанты) встречаются значительно меньше, чем родительские фенотипы.
Частота рекомбинаций (с) подсчитывается по формуле:
с = ((k + m) : n) × 100 % ,
где к, m – количество рекомбинантов,
n – общее количество потомков.
При скрещивании дигетерозиготной самки с серым телом и длинными крыльями с гомозиготным самцом с черным телом и короткими крыльями было получено в потомстве:
мушек с серым телом и длинными крыльями – 965;
мушек с черным телом и короткими крыльями – 944;
мушек с серым телом и короткими крыльями – 185;
мушек с черным телом и длинными крыльями – 206.
То есть можем подсчитать частоту рекомбинаций:
((965 + 944) : (965 + 944 + 185 + 206)) × 100 % = 17 % .
Частота рекомбинаций (частота скрещения хромосом) является косвенным отображением силы сцепления генов в хромосомах. Чем она больше, тем меньше сила сцепления генов в хромосоме и тем дальше гены локализованы один от другого. Тем больше возможность отрыва хромосом и больше возможность образования рекомбинантных особей. Частота кроссинговера может достичь 50 % , но не превышает этой величины. Это невозможно потому, что при очень больших расстояниях между генами возникает дополнительный обмен в других участках (двойной, тройной кроссинговер), часть кроссоверных форм может оставаться неучтенной, что в свою очередь приведет к общему уменьшению процента кроссинговера.
Если расстояние между хромосомами равно 50 % и больше – состояния признаков наследуются независимо.
Генетические карты хромосом
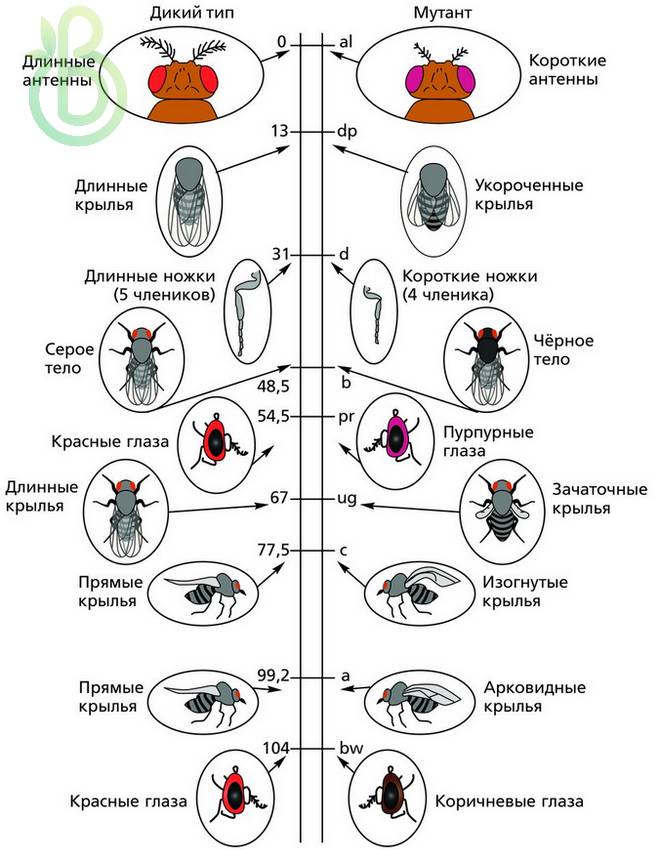
Знание частоты рекомбинаций дает возможность составлять карты относительного расположения генов в хромосомах. На генетических картах гены расположены линейно один за другим на определенном расстоянии, которое определяется в процентах кроссинговера (частота рекомбинаций) или в морганидах (1 % кроссинговера равен одной морганиде). Чтобы построить генетическую карту растений или животных, проводят анализирующее скрещивание, где достаточно подсчитать процент особей, которые образовались вследствие кроссинговера. Необходимо также определить число групп сцепления и принадлежность генов к ним.
Например, частота рекомбинаций между генами равна: А и В – 8%, В и С – 6%. Этих данных недостаточно для составления карты, так как возможны варианты.
Необходимо знать расстояние между генами А и С.
Для человека невозможно применение метода скрещивания (гибридологического), поэтому его заменили методом анализа родословных.
Генетическая карта хромосом – это схема относительного размещения генов в одной хромосоме, которые принадлежат к одной группе сцепления.
Первые генетические карты были составлены для дрозофил, а потом и для других объектов.
Генетические карты составляют для каждой пары гомологичных хромосом. Каждая группа сцепления имеет свой порядковый номер (римскими цифрами) в зависимости от порядка открытия. Кроме номера в каждой группе сцепления указывается полное или сокращенное название генов, расстояние каждого гена в единицах кроссинговера от одного из концов хромосомы, место нахождения центромеры.
В 1930 году был разработан новый способ картирования хромосом, основанный на наличии в слюнных железах насекомых больших хромосом – политенных, имеющих хроматин в виде чередования темных и светлых дисков при окраске, которые видно даже при небольшом увеличении микроскопа. При выявлении дефектов в темных хроматиновых дисках сравнивали их с изменениями соответственно локализованных генов. Такие карты стали называть хромосомными (цитологическими). Генетические карты сравнивают с цитологическими. Цитологические карты хромосом определяют хромосому как физическое тело. Порядок генов в этих картах идентичен генетическим. Таким образом, генетические карты указывают на реальный порядок генов в хромосоме. Расстояние между генами на генетической карте приблизительно, так как нет соответствия возле центромеры – на генетической карте гены расположены плотнее, чем на хромосомной. Это связано со сниженной частотой кроссинговера возле центромеры. В других участках хромосомы наблюдается соответствие.
При картировании генов человека, кроме анализа родословных, используют другие методы. Одним из них является метод гибридизации соматических клеток грызунов и человека в культуре ткани. В гибридных клетках при размножении теряются одна или несколько хромосом изучаемого вида. После анализа большого количества клонов клеток отбирают 20-30, которые отличаются по набору хромосом. Если у всех клонов отсутствует хромосома и отсутствует изучаемый белок, а у других клонов есть белок и хромосома, делают вывод: синтез этого белка связан с этой хромосомой. Использование этого метода позволило за короткий срок составить генетические карты человека и животных.
Благодаря успехам в молекулярной генетике используют еще один метод картирования генов. ДНК гена выделяют с применением методов генной инженерии. Наносят раствор этих генов с меченной (радиоактивной или флуоресцентной) последовательностью ДНК на митотические хромосомы. Эти участки ДНК включаются в хромосомах на «свое» место, которое можно определить.
В 2003 году была выполнена программа «Геном человека» (запланированная ООН): расшифрован геном человека, изучены все 24 группы сцепления.
Для составления генетических карт прокариот используют другие методы, которые связаны с особенностями их строения. Прокариоты – гаплоидные организмы. Составление генетических карт прокариот строится на существовании особого процесса у бактерий – конъюгации.
Важность составления генетических карт заключается в использовании их в селекции растений, животных и микроорганизмов. Генетические карты человека могут быть полезными в развитии медицины и здравоохранения для диагностики, предотвращения и лечения трудных наследственных заболеваний.
Хромосомная теория наследственности
Открытие сцепленного наследования, его нарушений позволило Т. X. Моргану создать хромосомную теорию наследственности. Она была дополнена современными фактами исследований генетики и цитологии.
Основные положения хромосомной теории наследственности:
– гены расположены в хромосомах, каждый ген имеет определенное место (локус) в хромосоме;
– гены в хромосомах расположены линейно;
– гены одной хромосомы образуют группу сцепления; количество групп сцепления равняется гаплоидному набору хромосом и постоянно для каждого вида;
– аллельные гены находятся в одинаковых локусах в гомологичных хромосомах;
– между гомологичными хромосомами возможен обмен аллельными генами (кроссинговер);
– расстояние между генами в хромосомах пропорционально проценту кроссинговера между ними; чем дальше гены один от другого, тем чаще между ними случается кроссинговер;
– гены относительно стабильны, но могут изменяться в результате мутационного процесса;
– каждый биологический вид имеет определенный набор хромосом (кариотип).
Взаимодействие неаллельных генов
Признаки большинства формируются с участием нескольких генов, взаимодействие между которыми отражается на проявлении фенотипа.
Типичные примеры взаимодействия аллельных генов – это полное доминирование, промежуточный характер наследования.
Но в процессе развития организма в сложные взаимодействия между собой вступают и неаллельные гены. Наиболее известные взаимодействия – комплементарность, эпистаз и полимерия.
Комплементарность
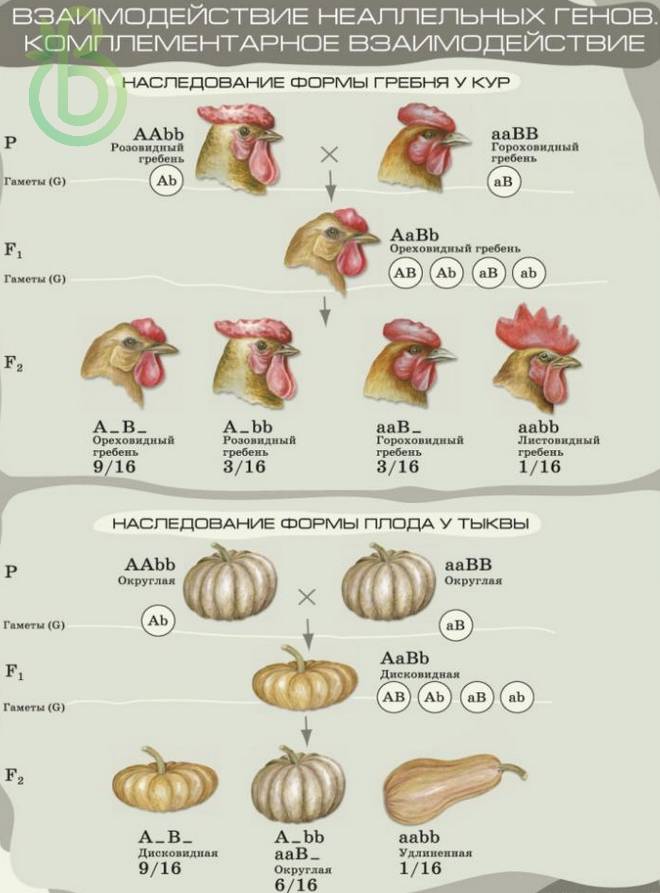
Комплементарные или дополнительно действующие гены – это таких два или более доминантных неаллельных гена, которые при взаимодействии определяют развитие нового признака. Например, два неаллельных доминантных гена горошка душистого предопределяют пурпурную окраску венчика, все другие соединения – белую. При скрещивании двух растений с белыми венчиками все у гибридов F1 растения были пурпурные. При самоопылении растений с F1 в F2 наблюдалось расщепление растений по окраске венчика в соотношении 9:7.
Эпистаз

Эпистаз (от греч. эпистазис – остановка, препятствие) – это такое взаимодействие неаллельных генов, когда аллель одного гена подавляет проявление аллели другого. Гены, подавляющие проявление действия других неаллельных генов, называются супрессорами. Они могут быть как доминантными, так и рецессивными.
Например, при скрещивании свиней черных и белых разных пород в F1 появляются белые. Скрещивание их между собой привело к возникновению белых (12/16), черных (3/16) и красных (1/16). Белые поросята имеют хоть один доминантный ген-супрессор I. Черные поросята гомозиготны по рецессивному аллелю i, что не препятствует формированию окраски, и несут доминантный аллель е, что способствует образованию черного пигмента. Красные поросята не имеют доминантных генов (генотип – eeii).
Полимерия
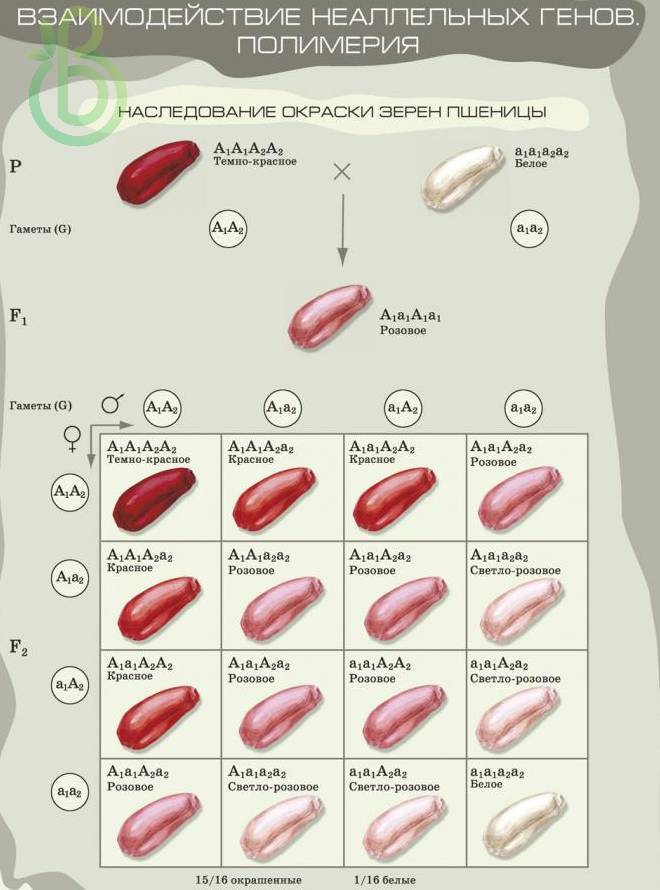
Полимерия (от греч. полимерия – многосложность) – это такое взаимодействие неаллельных генов, когда разные доминантные гены влияют на степень развития определенного состояния признака. Обозначают эти гены одной буквой с разными индексами. Чем больше доминантных генов в генотипе, тем больше степень проявления признака. Например, окраска зерна пшеницы может варьировать от белой до темно-красной. Растение, которое гомозиготно по рецессивным признакам (а1а1а2а2), будет иметь белую окраску семян. Хоть один доминантный ген будет определять окраску семян. Растения, гомозиготные по доминантным признакам (A1А1А2А2), будут иметь темно-красную окраску семян.
Множественное действие генов
Существует такое явление, когда на проявление состояний разных признаков влияет одна аллель. Называется оно множественным действием аллелей. Например, при заболевании человека арахнодактилией (человек имеет удлиненные пальцы конечностей, похожие на конечности паука) наблюдаются пороки сердца и неправильное положение хрусталика глаза. Арахнодактилия обусловлена мутацией доминантной аллели. Заболевание галактоземеем связано с рецессивной мутацией гена, который кодирует фермент, необходимый для усвоения клетками галактозы (молочного сахара). Вместе с заболеванием у людей развивается полоумие, цирроз печени, слепота.